När Sveriges Ornitologiska Förening i juni 1950 stod som värd för den tionde internationella ornitologkongressen, hade man under den andra kongressexkursionen till Abisko, turen att finna inte mindre än tio nordsångare sjungande utmed Nuoljas sluttning mellan Abisko, och Björkliden. Att detta var en sensation även för de svenska exkursionsledarna, framgår av Lundevalls (1951, 1952) entusiastiska och trevliga skildringar. Själv hade jag aldrig sett fågeln, när jag i juli 1952 på grund av ett särskilt uppdrag reste norrut för att söka få personlig kontakt med arten. Följande skildring är alltså huvudsakligen baserad på det ytterst begränsade studium, som kunde göras under drygt fyra dygn efter det att ett bo påträffats på Nuoljas nordsluttning på morgonen den 22 juli.
Nordsångaren hör alltjämt till de minst kända arterna i vår fauna, men åtskilligt har dock under årens lopp skrivits om den i Skandinavien. Tidigare utförligt och lättillgängligt beskrivna detaljer, såsom biotop, sång o.s.v., komma därför att i det följande behandlas mycket kortfattat, huvudsakligen med hänvisningar. Till en början må påminnas om Erik Rosenberg (1942) mångsidiga skildring av fågeln i Våra Fåglar i Norden.
Inledningsvis bör framhållas, att den skandinaviska populationen liksom huvudmassan av världens övriga nordsångare tillhör nominatrasen, Phylloscopus b. borealis. Nedanstående skildring bör alltså anses hänföra sig till denna ras. Om de subtila skillnaderna mellan denna och övriga raser se Ticehurst (1938) utomordentliga monografi över släktet Phylloscopus.

Fig. 01 Nordsångare hanen. Bilden visar tre artkaraktärer på vingen:
dels den ytterst korta första handpennan, som är knappt så lång som
vingtäckarna, dels det förhållandet att den andra handpennan är ungefär
lika lång som den femte, dels avsaknaden av inskärning på sjätte handpennans
ytterfan (däremot synes tydligt inskärningen på pennorna nr 3, 4, 5). Ingen
annan Phylloscopusart har dessa kännemärken i kombination med ett enkelt
vingband. Lundsångaren, Ph. trochiloides, har längre penna nr 1 och tydligt
kortare nr 2, medan nr 3, 4 och 5 äro inbördes liklånga och nr 6 nära
spetsen har inskärning på ytterfanet. (![]() Phylloscopus borealis male. Wing
characters: No other Phylloscopus but borealis has such a short first primary
and a single wing bar combined with the lack of emargination on the outer web of
the sixth primary.)
Phylloscopus borealis male. Wing
characters: No other Phylloscopus but borealis has such a short first primary
and a single wing bar combined with the lack of emargination on the outer web of
the sixth primary.)
Fältkännetecken. Nordsångaren har karakteriserats som mycket lik lövsångaren, Phylloscopus trochilus. Detta är naturligtvis riktigt. Emellertid är problemet för oss skandinaver närmast att skilja arten från lövsångaren. Det kan därför anmärkas, att fågelns allmänna gestalt för mig personligen synes mera lik grönsångarens, Phylloscopus sibilatrix, än lövsångarens, även om den i storlek kommer närmast den sistnämnda. I färghänseende såg särskilt min nordsångarehane närmast ut som en grönsångare med urblekt undersida.

Fig. 2. Nordsångaren landar vid boet med larver i näbben. Av bilden
framgår relationen i längd mellan handpennorna nr 2 - 6. (![]() Phylloscopus
borealis with a beak full of green larvae Wing characters.)
Phylloscopus
borealis with a beak full of green larvae Wing characters.)
Det distinkta dräktkännetecknet i fält, då man jämför med våra övriga Phylloscopus - arter, är som bekant det smala, ljusa tvärband över vingen, vilket bildas av de större vingtäckarnas gråvita spetsar. Emellertid kan detta under våren och sommaren helt nötas av. Så var fallet med honan i mitt nordsångarpar (se fig. 7.) Frånvaron av vingbandet är alltså icke något ovillkorligt indicium mot arten borealis. Det talas stundom om "två tvärställda vingband" men Tichhurst (1938:123), som granskat skinnen av 478 nordsångare av rasen borealis, säger att man blott tillfälligtvis kan finna en antydan till ett extra vitt band, bildat av spetsarna av de mellersta vingtäckarna ("occasionally a trace on median coverts"). Vi göra nog klokt i att följa Tichhurst i det fallet och tala om ett vingband ("single wing bar").
Nordsångare utan synligt vingband böra i fält vara mycket svåra att identifiera utan hjälp av lätena. Dessa kan man också få ta till hjälp, då det, t. ex. i södra Sverige, gäller att skilja nordsångaren från andra Phylloscopus arter med smalt, ljust vingband, närmast Ph. trochiloides, som iakttogs i Lund 1939 (Rudebeck 1942). Nordsångaren är emellertid som en storvuxen lövsångare, medan lundsångaren är spensligare än de, flesta lövsångare.
Eftersom jag hade den sällsynta förmånen att i ett och ett halvt dygn kunna iakttaga fåglarna på nittio centimeters håll, må här nedan färgbeskrivning lämnas:
Hanen. Ryggen, den hoplagda vingen och stjärten bildade i färghänseende en enhet med markerat gulaktig olivgrön färg på gråaktig botten. Det markerade strecket över ögat mycket ljust men med tydligt svavelgul ton. Kinder gröngult gråvita. Vingbandet ljust gråvitt. Undersidan ljust gråvit med en på 90 cm avstånd nätt och jämnt märkbar anstrykning av svavelgult. Hanen som helhet ljusare, dessutom större och grövre byggd än honan.
Honan hade hjässa och rygg grå med tämligen svag ton av olivgrönt, vingar och stjärt med mera markerat olivgrön ton. Bandet över ögat gråvitt med ytterst svagt svavelgul ton. Kinder svagt svavelgult gråvita. Undersidan ljust gråvit utan märkbar gul ton.
Ungarna vid ca 10 dagars ålder hade hjässa – rygg - stjärt starkt olivgrönt grå. Strupen gul, hela undersidan för övrigt gulvit.
Vingformeln är ett utomordentligt viktigt medel att skilja olika Phylloscopusarter med, n. b. om man har dem i handen. Den är noga beskriven i alla mera utförliga handböcker och nämnes här närmast därför att den ej tidigare illustrerats i svensk litteratur. Såsom fotografierna visa, är den första, "förkrympta" handpennan ej längre än vingtäckarna, medan handpennan nr 2 är av ungefär samma längd som nr 5. Hos lundsångaren däremot är nr 1 hela 7 - 12 mm längre än vingtäckarna och nr 2 är mycket kortare än nr 5.
Mjukdelarnas färg hos de vuxna: Näbbryggen brunaktigt svart, övernäbbens kanter hornfärgade. Undernäbben ljust hornfärgad utmed kanterna, på själva spetsen och runt om vid basen, men strax framför mitten var undersidan svartaktig hos hanen, svartbrun hos honan. Ben och fötter gråaktigt ljusbruna. Iris mörkt svartbrun. Praktiskt taget samma beskrivning har lämnats av J. A. Palmén (1887), som tillägger, att tårnas sulor äro "smutsigt citrongula", men den förtjänar upprepas, eftersom senare även de bästa handböcker så att säga förenklat beskrivningen.
Ungarna vid 8 - 10 dagars ålder hade näbbens yttersida gråsvart med gul övernäbbspets. Näbbvinklar svavelgula. Gapet kraftigt färgat, mättat orangegult.
Svenska fynd
Svenska häckningsfynd äro hittills blott tre och dessa äro belägna i två huvudområden inom Torne lappmark. Uttryckssättet är kanske vilseledande - meningen är att framhålla, att fynden blivit koncentrerade till två huvudområden, men därmed är inte sagt, att fågelns förekomst i Sverige skulle vara uppdelad på och begränsad till just dessa områden.
Det första bofyndet gjorde Sune Hederström 1916 vid Sevujoki norr om Jukkasjärvi (Hederström 1944). Omkring 15 kilometer norr om denna plats fann Rosenberg (1942) fågeln 1939, 1941 och 1942 i Vittangijärvi området under sådana omständigheter. att det är troligt att den häckat där. Någon halvmil söder om Rosenbergs lokaler fann Mauritz Magnusson (in litt.) en sjungande hane den 28 juni 1950.
Själv genomsökte jag den 1 - 2 juli 1952 med hjälp av tre energiska medhjälpare (se uppsatsens slutord) så gott som varje kvadratmeter av de kända nordsångarbiotoperna vid Vittangijärvi med det resultatet, att det syntes osannolikt, att fågeln bodde där det året. Om nordsångare skulle ha funnits där under 1952, borde de dels ha anlänt före vårt besök (jfr nedan sid. 63), dels ha varit i sångtagen då. Kai Curry - Lindahl hörde dock den 3 juli i högre belägen och torrare björkskog en Phylloscopus sjunga på ett sätt som gör det troligt att det var en borealis.
Kring Torne träsks västända ha under senare år en anmärkningsvärd rad observationer gjorts. Det andra svenska bofyndet gjorde Lennart Andersson (in litt.) den 26 eller 27 juli 1940 omkring 15 km VNV Torneträsk, närmare bestämt i en bäckravin mellan riksrösena 268 A och 269. Fyndet är omnämnt av Rosenius (Svenska Fåglar och Fågelbon vol. 6:223), men på grund av ett tryckfel har det förefallit vara på norskt område. Det gjordes emellertid då Andersson med en radiogränspatrull låg på svenska sidan om gränsen mot Norge. Däremot söktes fågeln förgäves i samma område 1942 och 1946 L Andersson in litt.).
På fjället Nuoljas fot i närheten av Abisko hörde Gunnar Gustafsson (munt. medd.) en nordsångare sjunga under ett par timmar på morgonen den 23 juni 1949. Eftersom hemresan anträddes samma dag, blev det ej tillfälle till vidare efterforskningar. Följande år inträffade den stora invasion av både nordsångare och ornitologer på Nuolja, vilken Lundevall (1951, 1952) ingående skildrat. Sista veckan i juni 1951 räknades 5 sjungande nordsångare på Nuoljas nordsluttning (Ulfstrand 1953) och en hördes den 27 juni 1951 av fru Elisabeth Täcklind 4,5 km S Abisko (Lundevall 1952), tydligen omkring ½ km från den plats, där Ulfstrand hörde en hane sjunga den 4 juli samma år.
Under sommaren 1952, som började med ovanligt ihållande ruskväder. uppehöllo sig däremot, av allt att döma, endast två nordsångarhanar på det numer så bekanta tillhållet mellan Abisko och Björkliden. Björn Holm iakttog den ena den 22 och båda den 27 juni, sista gången även en hona. Av beskrivningen att döma var det just dessa fåglar jämte ytterligare en hona engelsmannen William Dady såg den 26 juni (W Dady in litt). Det ena parets bo påträffades, som ovan nämnts, den 22 juli. Dadys iakttagelse, att även den andra hanen hade en hona i sällskap, i kombination med den absoluta överensstämmelsen i de båda hanarnas sångbeteende under mina dagar på Nuolja, gör det ytterst troligt, att även paret nr 2 hade bo på fjällsluttningen 1952.
Utom de ovan nämnda ha några mera isolerade fynd gjorts norr om 68 breddgraden (jfr Holm 1950), och två gånger har David Sjölander (1946: 102) funnit den i Sjaunja området i Lule lappmark.
Kontentan av sammanställningen ovan blir den, att nordsångaren synes vara en årsviss men sparsamt förekommande häckfågel i Torne lappmark och att förekomsten fluktuerar ganska starkt, en sak som förefaller naturlig i denna yttersta utpost av artens häckningsområde i världen. Det är anmärkningsvärt, att den på ett av naturvetare så pass flitigt frekventerat fjäll som Nuolja ej påträffats förrän på 1940 - talet. Men sångperioden är kort och inträffar, när den ornitologiska högsäsongen är förbi, så att det vore måhända väl djärvt att nu draga vissa slutsatser om tidsförloppet för artens invandring i vårt land. I Norge var nordsångaren ännu 1875 okänd, men när Collett (1877: 10 - 11) 1876 reste till Finnmarken "med specielt Hensyn paa de østlige Sylvieformer" fann han med en gång "Ph. borealis ... udbredt over en stor Del af Finmarken", närmare bestämt över ett område som sträcker sig 400 - 450 km i öst - västlig riktning. Redan därav kan man förstå, att den näppeligen var nyinvandrad just då. Än mera talande är det, att arten ännu efter 70 år har i stora drag samma utbredning i Norge (Løvenskiold 1947 - 49).
Men de, som ha tillfälle att söka fågel i björkskogen i norra Lappland kring månadsskiftet juni - juli, böra hjälpa oss att vidga (dess kända utbredningsområde i Sverige!
I nordligaste Norge finns nordsångaren, som nyss antyddes, från västsidan av Porsangerfjorden österut genom Finnmarken. I Finland finns den ned till 68 breddgraden Finnilä 1918, Hortling 1929 - 31), i europeiska Ryssland ned till ungefär i jämnhöjd med Onega.
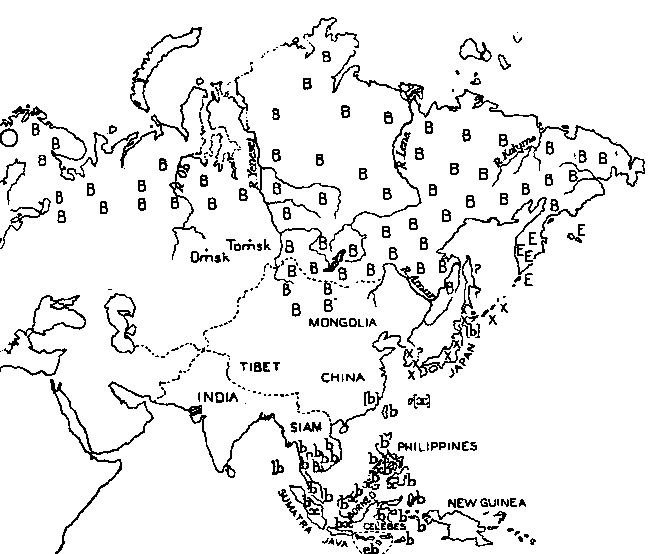
Fig. 3.
Nordsångarens utbredning i Gamla Världen (efter Ticehurst 1938, med
tillfogande av det svenska förekomstområdet). Phylloscopus b. borealis:
häckningsområde B. vinterfynd b, svenskt område 0. Phylloscopus b.
xanthodryas: häckningsområde X. vinterfynd x. Phylloscopus b. examinandus:
häckningsområde E, vinterfynd e. Phylloscopus b. kennicotti häckningsområde
Alaska, strax utanför kartans östra vintertillhåll se sid. 74. (![]() Breeding and
winter ranges of Phylloscopus borealis from Ticehurst 1938 Swedish breeding area
added by a circle)
Breeding and
winter ranges of Phylloscopus borealis from Ticehurst 1938 Swedish breeding area
added by a circle)
I Asiens norra hälft har nominatrasen ett mäktigt utbredningsområde, vars utsträckning framgår av kartan, vilken hämtats från Ticehurst (1938). Den finns där så längt i norr, som björkskog sträcker sig, d. v. s. till 75o N på Taimyrhalvön, och i söder ända ned till Khangai - bergen i Mongoliet (Ticehurst) det vill säga omkring 48o N. Detta är ungefär samma breddgrad som för Wien eller Paris, vilket måhända förefaller märkligt för vår nordsångare, men den har anledning, att känna sig hemma där, ty det är inom regioner med ständigt frusen mark, och den alpina faunan i Trans - Baikal hyser för oss så välbekanta arter som fjällripa, fjällpipare och berglärka (Berg 1950: 310).
Längst i öster ersättes rasen borealis av några andra. Den ena av dem är den större Phylloscopus borealis examinandus, som hör hemma på Kamtschatka och de nordligaste Kurilerna omedelbart söder därom (Bergman 1935).
I Japan och på de sydligaste Kurilerna har man rasen Ph. b. xanthodryas, vilken också skall ha ett genomsnittligt vingmått, som ligger högre än det som gäller för Ph. b. borealis. Xanthodryas är dessutom mera grön och mindre grå på ryggen, medan undersidan är mera gultonad än hos Ph. b. borealis. (Ticehurst).
1948 funno A. A. Allen och H. C. Kyllingstad i västra Alaska den första kända häckningsplatsen för Numenius tahitiensis (vars amerikanska namn är the bristle - thighed curlew", d. v. s. "spoven med borst på låren"). Det hette då, att nu hade amerikanerna löst sin sista stora ornitologiska gåta, sedan J. D. Soper 1929 gjorde det första fyndet av en boplats tillhörande den million blågäss, som man varje vinter sett överflyga USA till vinteruppehållet i och vid Missisippis delta. Men det finns massor av andra olösta problem, ej minst i det ofantliga Alaska, mot vilket vårt inre Lappland ter sig ganska lättillgängligt. Phylloscopus borealis är Amerikas enda bofasta representant för familjen Sylviidae, men ingen har ännu funnit dess bo i västra Alaska, där den skall förekomma. Joseph Dixon (1938) säger visserligen att den i juni 1926 var en "tämligen vanlig häckfågel vid övre delen av Savage River" i Mount McKinley National Park. Men han fann inget bo då och sökte arten förgäves vid upprepade besök på exakt samma lokaler sommaren 1932. De relativt få exemplar, man insamlat, synas visa, att Alaska - nordsångarna äro en särskild ras med mindre vingmått än övriga borealis och gulare undersida, varför denna ras fått namnet Ph. b. kennicotti. Den måste emellertid studeras betydligt mera på ett större material än hittills, innan dess hållbarhet kan garanteras och dess utbredningsområde avgränsas, säger Ticehurst (1938: 132). Tucker (1949: 332) ifrågasätter om det inte här är fråga om en alltjämt pågående kolonisering av Alaska, utgående från angränsande delar av Asien. K. C. Parkes & D. Amadon (1948) betrakta kennicotti som särskild ras.

Fig. 4. Nuoljas skogsgräns den 22 juli 1952. Nordsångarens boplats låg i
översta kanten av den sammanhängande björkskogen nära bildens mitt. (![]() Phylloscopus
borealis Nest site was among the uppermost birch trees on the slope July 1952.)
Phylloscopus
borealis Nest site was among the uppermost birch trees on the slope July 1952.)
Alla skandinaviska författare ha varit överens om nordsångarens biotopkrav: högvuxen björkskog med frodig undervegetation, gärna i närheten av någon bäck eller försumpade skogspartier. Boet lägges dock på någon torr plats i skogen (Finnilä 1918). Kanske kan en alltför snäv tolkning av nordsångarens biotopkrav i enstaka fall ha försvarat försöken att finna konkreta häckningsbevis. För mig personligen var boets läge på Nuolja 1952 en överraskning. Det låg i björkskogens övre kant på fjällets östsluttning, där det är så brant, att jag, när jag nalkades nedifrån, måste klättra de sista tio meterna mer eller mindre på alla fyra. Ur anteckningarna, gjorda på platsen, citeras: "Ett tunt torvjordskikt täcker fjällets klippgrund, men en och annan naken klippvägg sticker fram. Björkarna stå glest och är knotiga, låga fjällbjörkar. Videbuskarna, Salix lanata m. fl., äro likaledes glesa och låga här. Utmed vissa klippkanter och lodräta moss - och Saxifraga aizoides - klädda jordväggar sipprar litet droppande vatten. Vegetationen i örtväg präglas av Saxifraga aizoides, Thalictrum alpinum, Bartsia, Viola billora Solidago virgaurea, Veronica alpina, Cassiope tetragona, Salix reticulata. Boet är beläget strax ovan topparna av de översta raka men glesa björkarna i den mera yppiga fjällvegetationen, men sådan biotop, som anses vara utmärkande för nordsångaren, börjar först något längre ned på sluttningen.
Boet har byggts i ett alldeles lodrätt avsnitt av branten i ett väldränerat torrt parti, lätt skyddat av överhängande tunna, glesbladiga videkvistar från en strax ovanför växande videbuske. Det ligger i liksom en djup grotta i jordväggen innanför en gles ridå av fem torra videkvistar, som hänga ned just för öppningen. Boplatsen skulle passa en ringtrast förträffligt!" Se fig. 4 o. 5.
Björn Holm som introducerade mig i nordsångarreviren på Nuolja och gav mig den noggranna beskrivning på var hanarna höllo till den 27 juni, utan vilken boet aldrig skulle blivit funnet, meddelade (in litt.), att de "endast högst sällan och då blott kort tid vistades på så hög nivå, som där boet senare fanns".
Till jämförelse må nämnas, att de först funna skandinaviska bona enligt Collett (1893 - 94) lågo på följande sätt: det första "ved Foden af en brat skovklaedt Li, opunder en Traerod, der tildels dannede Redets Tag", det andra "i en liden Bakke mellem höie Traer och det tredje "i en høi og stärkt Skraanende Birkeli". Hederström och McNeile (1952) funno 1949 - 51 fem bon i Sydvaranger, alla placerade i sidan av mossklädda tuvor i tydligen låglänt, fuktig björkskog.
För att ej lämna en ensidig bild av fågelns fordringar på biotop bör erinras om att den särskilt i Asien även anträffas i barrskog. Taczanowski (1872: 358) uppger t. ex. ur Dybowskis anteckningar från östra Sibirien, närmare bestämt områdena kring Baikalsjön: "Till i mitten av juni uppehålla de (nordsångarna) sig i dalarnas björkskogar ... Vid mitten av juni uppdela de sig i par, lämna dalarna och bege sig högre upp för att häcka. De bo i mörk skog av cembratall … i många trakter gå de ända upp till skogsgränsen. De älska särskilt fuktiga, mörka ställen, där marken runt omkring är bevuxen med Rhododendron chrysanthum och Salix" Och Stegmann(1931: 192) skriver om nordsångaren i östra Sibirien: "... den bebodde huvudsakligen skogsbältets övre gräns och den subalpina zonen. Vanligast fann jag den vara i kryptallarnas snårskog ... I Pinus pumila - zonen voro ungarna ännu den 26 juli i sina bon."

Fig. 5. Nordsångarens, bo med ca 7 dagar gamla ungar den 22 juli 1952.
Observera hur väl det är inbyggt i det vertikala marktäcket. Två av de fem
torra videkvistar, som hängde ned för öppningen, ha avlägsnats före
fotograferingen, likaså åtskilligt grönt. Vad man såg av boet gjorde mest
intryck av ett klot av gulgrön mossa. (![]() Phylloscopus borealis, nest with young,
approx. 7 days old. As seen nest was built for the main part of moss.)
Phylloscopus borealis, nest with young,
approx. 7 days old. As seen nest was built for the main part of moss.)


Fig. 7. Nordsångare hona fotograferad under den enda halvtimme på dagen,
då solen kunde nå boet, kl. 5 på morgonen den 26 juli 1952. Inget ljust
vingband! (![]()
Fig. 8
. En unge har fått sin mat och gjort helomvändning. Nordsångarehonan griper i nästa ögonblick en spillningsboll, innan denna hinner ramla i boet. 26 juli 1952. (

Fig. 9. En situation motsvarande den på föregående bild. Här ser man
fågelns väl markerade näbborst. (![]() Phylloscopus borealis, as fig. 8. Rictal
bristles are seen.)
Phylloscopus borealis, as fig. 8. Rictal
bristles are seen.)
a) Sången har tydligen fängslat de flesta iakttagare och den har beskrivits av många författare, särskilt väl av Schaanning (1907), Rosenberg (1942) och Lundevall (1951, 1952). Det är därför onödigt att ånyo komma med en utförlig beskrivning.
När jag själv första gången på 150 - 200 meters avstånd fick höra den för mig då obekanta sången, anade jag visserligen strax, vem det var. Men samtidigt tvangs jag att i hast tänka över frågan: om fågeln skulle ha tystnat och försvunnit, innan jag hinner fram, vilka andra alternativ skulle då kunna tänkas? Med tanke på att det var björkskog vid Torne Träsk, kunde visserligen ingen annan art rimligtvis tänkas, men det slog mig strax, att de 5 - 7 stavelserna i varje tonräcka, tillillillillillill kommo med tjockt I-ljud" rullande på ett sätt som starkt påminde om entitans enkla sång, ehuru tonerna kommo snabbare, i själva verket avsevärt snabbare än man kan förstå av den för mänskliga röstorgan tillrättalagda bokstaveringen ovan. Detta gällde båda hanarna. Efter hemkomsten från fjällen fann jag, att även Rosenberg talat om likheten med entita, och med en travestering av Lundevalls uttryckssätt, skulle jag vilja säga, att sången låter som om den kom från en grönsångare som försöker sjunga som en entita.
Mest av allt liknar nordsångarens sång visserligen berglövsångarens, Ph. bonelli, vilket flera kongressdeltagare påpekade vid besöket på Nuolja 1950. Men denna liknelse ha de flesta av oss svenskar föga glädj e av!
Erik Rosenberg (1942) säger att man hör sången från strax före midnatt till sent på eftermiddagen. Och Sten Bergman (1935) säger om Ph. b. examinandus på Kamtschatka, att han särskilt lade märke till "att denna lilla fågel inte lät sig störas det ringaste av vädret. När det var regnväder och alla andra fåglar tystnat, hörde man dock fortfarande nordsångarens sång."
Det var sålunda intressant att konstatera, att våra två banar på Nuolja 1952 totalt upphört att sjunga under tiden 22 - 25 juli, då jag nästan hela dagarna vistades i deras tillhåll och min personliga sovperiod i stort sett torde ha sammanfallit med deras. Av detta kan man draga slutsatsen, att nordsångaren i likhet med så många andra släktingar upphör att sjunga, sedan äggen kläckts. Samma erfarenhet gjorde Collett (1893 - 94) i Finnmarken. Denna vana ökar givetvis i hög grad svårigheterna att finna boet.
En överraskning väntade mig sista morgonen på Nuolja. Klockan 01.30 - 02.45 den 26 juli var jag i området för nordsångaren nr 2 utan att höra något. Men vid min återkomst klockan 03.45 sjöng denna hane, och när jag en stund senare kom bort till nr 1, hade även den hanen börjat sjunga och han fortsatte med det nästan oavbrutet en hel timme. Därefter omväxlade han med sång och matning av ungarna, tills jag definitivt avlägsnade mig kl. 8.
Den förklaring, som förefaller troligast, är den, att båda hanarna nu nått ett sådant stadium i häckningsperioden, då de åter börja en kort sångperiod. I det kända boet voro ungarna ca II dagar gamla, alltså redo att lämna det vilken dag som helst.
Man kan fråga sig, om inte väderleken kunde ha spelat en viss roll. För att undersöka detta vände jag mig till föreståndaren för Abisko Naturvetenskapliga station, Gustaf Sandberg, och fick termo - och barogram för tiden 22 - 26 juli. Av dessa kan man emellertid vid jämförelse mellan de olika dagarna inte spåra någon trolig påverkan från vädrets sida.
b) Orosläte. "De gamle fugle tilkjendegav sin aengstelse her ved et skrig, der fuldstaendig lignede lokketonen hos Cinclus ... ". säger Schaanning (1907: 19). Mina egna anteckningar säga:"Varningslätet är ett korthugget "dsri", som särskilt hos hona är närmast likt gulsparvens lockton. Även hos hane är lätet närmast gulsparvlikt men hos honom har det en stark dragning åt strömstare - lockläte. Hanens läte är nämligen skarpare, honans mjukare". Schanning hade bokstaverat lätet som "sir!" I själva verket kommer r'et varken före eller efter "i" utan det anger ett surrande som ligger i själva vokalljudet och detta surrande förefaller mig markeras bäst genom att sätta "r" före "i". Med denna bokstavering får man också bättre fram lätets korthuggna karaktär.
Av intresse är, att detta läte används på två helt olika sätt. När jag uppehöll mig i närheten av boet, upprepades det i ganska långsam takt - det fungerade då uppenbarligen som varningsläte med uppgift att ge ungarna impulsen att hålla sig stilla, resp. att väcka den andra vuxna fågelns uppmärksamhet. När man satt gömd invid boet kunde man iakttaga, hur ungarna prompt reagerade med stillhet, även om varningslätet var rätt långt borta.
Men när man från fjällsluttningen nedom boet hörde trädgårdssångare och andra fåglar träta så, som om en hermelin eller katt vore i farten, hördes nordsångaren gå på i samma häftiga takt som en svartvit flugsnappare (Muscicapa hypoleuca) i samma situation. Då användes samma läte som »vredesläte» av den typ, som, för att tala med Konrad Lorenz, har till funktion att göra situationen otrevlig för en inkräktare.
Lockläte. Ett läte, som verkligen tycktes fungera som egentligt lockläte, eftersom honan ibland lät höra det, när hon utan oro nalkade, boet och ungarna därvid inte tryckte, var en ytterligare förmjukad upplaga av ovannämnda dsri Det bokstaverades »dsré».
När de 10 – 11 dagar gamla ungarna lågo i lugn och ro i boet och blott pysslade med sig själva, läto de ibland höra ett läte, som på mycket nära håll kunde liknas vid en svag upplaga av den svartvita flugsnapparens varnings - »hyi», alltså inte lövsångarens mjuka läte av samma bokstavering. Vilken roll, detta läte spelar, är ovisst - det föreföll närmast vara ett lockläte. Möjligen är detta läte detsamma som det »tswee - ip» (engelskt uttal!), vilket Tucker uppger, att man hört från nordsångare på flyttning.
Andra läten ha i förbigående beskrivits av Finnilä (1918), Tucker (Witherby 1945) m. fl.
Ungarnas skräckläten och - attityd.
När man rör sig omedelbart intill boet med 9 - 11 dagar gamla ungar eller när man petar i ris etc. helt nära, reagera ungarna med en sorts skräckbeteende: de göra diminutiva symboliska utfall, medan de göra hastigt klappande rörelser med gapet under ett knäppande ljud, som låter som om det uppstode genom att näbben slås ihop. Samtidigt hör man ett fräsande eller surrande ljud. Det hela påminner osökt om en fjällämmel som sätter sig till ilsket motvärn, när han blir trängd.
Beteendet känner man även från andra Phylloscopus - arter. Jouard (1935:129) har på grundval av sina egna erfarenheter av berglövsångaren, Phylloscopus bonelli, ifrågasatt, om det verkligen är ett skräckläte, och vill tro, att det är ett normalt »matningsläte». Så är det icke för nordsångaren. När dennas ungar matas, hör man på nära håll ett ivrigt sirrande ljud, ej särskilt starkt, vilket är av helt annan natur än »skräcklätet».
Målmedvetna rovdjur lära dock knappast bli skrämda av fågelungarnas små »utfall». Däremot är det tänkbart, att det nämnda beteendets uppgift skulle kunna vara att motverka skadegörelse från icke rovdjur, t. ex. hindra betande djur att sticka mulen i boet.
Nordsångaren anländer till Sverige långt senare än alla andra flyttfåglar. Grå flugsnapparen, Muscicapa striata som hör till de allra senaste i södra Sverige, anländer till norra Lappland o. s. v. i början av juni, t. ex. Laisdalen vid Svaipa 3.6.1948, Pajala - trakten 7.6.1951, övre Lainio 10.6. eller tidigare 1949. Och trädgårdssångaren, Sylvia borin, kom till Laisdalen vid Svaipa den 6.6.1948, till Vindeldalen 15.6.1944, till Abiskojaure 17.6.1939. Men nordsångaren är i Sverige tidigast iakttagen den 16 juni 1950 (B. Holm 1952), till Vittangijärvi har den kommit dagarna före midsommar (Rosenberg in litt.) och enligt Lundevall (1952) skulle hela kontingenten på Nuolja 1950 ha anlänt natten till den 26 juni, alltså först tio dagar efter det att Holm hörde den sjunga vid Nakerijärvi. Vid Holms besök pa Nuolja den 22 juni 1952 hördes blott en nordsångare sjunga.
Till jämförelse bör nämnas kända ankomsttider i Norska Finnmarken. Tidigaste datum är 12.6. 1897 (Wessel), 13.6.1908 (Schanning 1926). 14.6. (Hederström), 18.6.1906 (Schanning 1907), och Collett (1893 - 94) nämner att under 1885 var den nyanländ den 22 och sjöng den 23, medan hona iakttogs först den 26 juni. Enligt Blair (1936) observerades den första den 17, ytterligare en den 18, medan många voro komna den 20 juni 1927. 1 Finland är den iakttagen den 19.6.1937 Ruthke och 19.6.1949 (T. Lampio enl. Orn. Fenn. 29:103).
Rosenberg (1942) säger, att nordsångaren anländer först då björklöven spruckit ut, och Holm (in litt.) delar den åsikten och säger att björklöven just börjat slå ut, då han såg fågeln den 16 juni 1950.
Som sammanfattning kan alltså sägas, att nordsångaren till sina boplatser i Norra Lappland vanligen anländer i sista hälften av juni.
Intressant är att jämföra med uppgifter från området vid Baikalsjön i södra Sibirien, där avståndet från boplatserna blott är tredjedelen så långt som för de skandinaviska nordsångarna. Dit anlända de i slutet av maj eller början av juni, men de uppehålla sig då i dalgångarnas björkskog och dela upp sig i par först vid mitten av juni, då de lämna dalarna och bege sig högre upp för att häcka (Taczanowski 1872).
Lundevall (1952:18) anger att lövsångaren under 1950 skulle lagt sitt första ägg först 30 - 32 dagar efter ankomsten till Abiskotrakten. (I England har MAY (1947: 6) iakttagit hur en lövsångarhona började bygga 5 dagar efter sin egen resp. 14 dagar efter första hanens ankomst på platsen, detta 1945. Året dessförinnan började bobygget 16 dagar efter hanens ankomst. Detta betyder att första ägget lades ungefär 3 veckor efter första hanens ankomst). Avsevärt snabbare är nordsångaren, vilket kan ses som en logisk följd av den sena ankomsttiden.
Boet på Nuolja 1952 hade den 22 juli sju stycken minst 7 dagar gamla ungar. Räknar man som en arbetshypotes med 12 - 13 dagars ruvningstid, skulle detta innebära, att sista ägget lagts den 2 – 3 juli. (För lövsångaren ange Niethammer och Witherby normalt 13 dagar, för grönsångaren anger v. Treuensfels 13, Steinfatt 12, 13, i något fall 14 dagar. Ruvningstiden för Phylloscopus bonelli är 13 dagar enl. Niethammer). Första ägget skulle lagts den 26 - 27 juni 1952. För lövsångaren anger Kuusisto (1941:31) en bobyggnadstid av 5 - 6 dagar. Nu hade Holm den 22 juni blott observerat en hane på det parti av Nuoljas sluttning, där han den 27 juni fann två hanar och en hona.
Denna och andra iakttagelser ge oss anledning till slutsatsen: Nordsångaren
synes börja bygga bo så gott som omedelbart efter honans ankomst. I boet vid
Jukkasjärvi 1916 måste första ägget ha lagts senast den 23 juni. I Lennart
Anderssons bo norr om Lapplandia "torde äggen ha blivit lagda omkring den
6 juli" (Rosenius, Vol. 6:228), vilket synes innebära, att första ägget
lagts omkring den 1 juli. Detta stämmer tämligen väl överens med uppgifter
för Sydvaranger, varifrån följande data om äggläggningens början finnes:
C:a 22 juni 1904 (en kull enl. Wessel 1905 - 06:32)
25 juni - 1 juli 1949, -50, -51. (för 5 olika kullar enl. McNeile 1952: 8 - 9)
C:a 1 juli. (Eglaegningen i förste uge af juli enl. Schanning 1907:19)
2 juli 1899 (en kull enl. Wessel 1905-06)
9, 10 resp. 11 juli 1885. (för tre olika kullar enl. Collett 1893-94: 21.)
Normalt börjar alltså äggläggningen i norra Skandinavien under sista veckan
av juni eller första veckan av juli.
Det har sitt intresse att konstatera, att Stegmann (1931:192) anger ungefär samma häckningstid för nordsångarna i östra Sibirien, trots att våra skandinaviska nordsångare torde ha cirka 3000 km längre väg att färdas från vintertillhållen. I skogen uppe i bergen väster om Okotska sjön voro ungarna just utflugna den 26 juli, medan ungarna i Pinus pumil - zonen vid den tiden ännu voro kvar i sina bon.
Uppgifterna om revirets omfång variera från »en cirkel med 10 - 25 m radie» till »1000 m i fyrkant». Av detta drar man snabbt och lätt slutsatsen, att vidare studier äro önskvärda, innan man uttalar sig om artens vanor beträffande revir i egentlig mening. Möjligen är det så, att hanarna på grund av populationens gleshet i våra trakter ofta röra sig långt utanför ett område, som motsvarar vad man menar med begreppet revir i strikt betydelse.
Betecknande torde emellertid - att döma av många olika litteraturgifter - vara Björn Holms uppgifter (in litt.) att hanarna på Nuolja 1952 vardera hade ett sång - tillhåll, som mätte omkring 300 meter i längd och 150 meter i Bredd. Men boet låg i utkanten av detta område och hanen sjöng sällan i närheten av boet.
Värdefull är också McNeiles (1952: 2) uppgift, att hanarna, när honan börjat ruvningen, sjöngo tämligen konstant inom 100 steg från boet, ehuru de ej alls gåvo någon antydan om i vilken riktning detta befann sig.
Den enda tillgängliga beskrivningen av parningsbeteendet har helt nyligen lämnats av Ulfstrand (1953: 27), och den återgives därför endast i det engelska sammandraget här nedan.
Tucker (Witherby 1945) har hört ett »sit-sit-sit-sit-», mera stackato än sången, under »sexual chase» och Hederström har hört en hane sjunga en mera melodiös version av sin vanliga sång, då honan slöt sig, till honom (McNeile 1952: 8).
Av litteraturen att döma bör boet på Nuolja 1952 ha varit ett ganska karakteristiskt nordsångarbo. Det var av den för släktet Phylloscopus utmärkande typen - ett ihåligt klot med en liten ingång från ena sidan. Det var helt av mossa och fina strån. Kupolen var av mossa, inåt klädd av ett ytterst tunt, glest men jämnt skikt av fina strån, som nedtill övergick i den även av strån bestående balen. Utåt såg man praktiskt taget endast mossa, vilket kom en att tänka på järnsparvens sätt att bygga sitt bo. Se bild 5. Löv ingick ej i bygget.
Finnilä (1918) säger sig i Finland ha funnit ett bo, som var byggt uteslutande av torrt gräs, men detta förefaller vara en mera ovanlig företeelse. Däremot tycks det vara vanligt att det är liksom inbyggt under botuvans eget mosstäcke.
Boets särprägel. Från löv - och gransångare skiljes boet genom att det helt saknar fjäder i balen. Från grönsångaren, som ej heller fodrar sitt bo med fjäder, torde det i de allra flesta fall skiljas genom sin mer eller mindre starka anhopning av mossa i hela kupolen. (Vid en undersökning av grönsångaren fann Steinfatt, 1937: 184, att samtliga 7 bon voro praktiskt taget uteslutande byggda av torrt gräs). Grönsångaren har ofta en påfallande anhopning av torra löv på kupolen, vilket torde vara sällsynt undantag för nordsångaren.
Beträffande fjäder i boet finnes verkligen en uppgift av Carpelan (1927) att han i ett bo i Finland påträffat tre små fjädrar, sannolikt av fjällvråk, i ett bo. Men detta synes vara ett helt isolerat undantag.) [En uppgift i ett stort amerikanskt verk, att enligt Riely (1918: 620) ett bo, fodrat med vit fjällripfjäder, skulle anträffats i östra Sibirien, är däremot felaktig. I detta, som i så många andra fall i litteraturen, har man förväxlat Ph. trochilus eversmanni med »Eversmann's warbler», som är det engelska namnet för Ph. b. Borealis]
Förhållandet att t. ex. löv - och gransångare alltid och vanligen rikligt fodra boet med fjäder, medan de närstående arterna grön -och nordsångare aldrig göra det, är ett gott exempel på hur fåglarnas instinkthandlingar löpa i stelt fastlåsta banor.
Efter denna reflexion kan man knappast lämna bobyggnadsbeskrivningen utan att kasta ett öga på hur vanan att fodra boet med fjäder respektive utan fjäder fördelar sig inom släktet Phylloscopus.
Man finner då, att det är en alldeles särskild grupp arter, som har vanan att icke använda fjäder. Det är de, som tidigare förts till ett särskilt undersläkte, Acanthopneuste, vilket emellertid underkändes av Hartert & Steinbacher (1932-38) och Ticehurst (1938: 10), I Bents stora verk, tryckt 1949, användes dock detta släktnamn för borealis, vilken f. ö. räknats just som typart för Acanthopneuste.
Om dessa arter och vissa morfologiska karaktärer hos dem säger Ticehurst »Jn all species which have been grouped in the genus Acanthopneuste the nasal hairs extend to or beyond the end of the nasal groove, viz. borealis, magnirostris, trochiloides, reguloides, davisoni, tenellipes, coronatus and occipitalis, to which I should add Cantator and ricketti. But this character does not of itself define the genus Acanthopneuste unless the charaeter of a wider and stouter bill - purely a comparative one - is added, as in maculipennis, and in sibilatrix the nasal hairs extend also to the end of the groove and nearly as far in bonelli. Even so, the genus Acanthopnuste is a very weak one and to be consistent another genus would have to bo used for sibilatrix with its bill of a Phylloscopus and nasal hairs of an Acanthopneuste!»
Hos dessa 13 arter, som antingen räknats till Acanthopneuste eller bära den viktigaste Acanthopneuste - karaktären, känna vi bobyggnadsvanorna för åtminstone 11 stycken och samtliga dessa fodra boet med strån eller hår eller i något fall (cantator) med växtull, men ingen av dem använder fjäder. (Uppgifter om undantag finnas för reguloides.) Ph. presbytes, endemisk stannfågel med en ras på vardera öarna Timor och Flores i den malajiska arkipelagen mellan Borneo och Australien, har »näshår», som en Acanthopneuste, men dess bo är okänt.
Av de övriga bilda 3 st. den s. k. Reguloides - gruppen, kungsfågellika sångare, och av dessa synas proregulus och subviridis använda fjäder, medan inornatus använder hår.
Återstår så ytterligare 13 arter, bland vilka vår vanliga lövsångare och gransångaren med sina åtta raser m. fl. befinna sig. Bobyggnadsvanorna äro kända för minst 9 av dem, och samtliga dessa använda fjäder. May (1947:7) har i ett lövsångarbo räknat 240 fjädrar!
Detta förhållande, att en markant, biologisk skillnad synes existera mellan arter med Acanthopneuste - karaktärer och övriga arter synes värt allt beaktande. Det tyder i utvecklingshistoriskt hänseende på att de förstnämnda arterna höra inbördes samman på ett djupare sått än de små morfologiska karaktärerna antyda.
Relativt få nordsångarägg äro studerade. Noggranna beskrivningar av vissa kullar finnas hos Hederström (1944), McNeile (1952) och Rosenius.
Att angiva en distinkt särprägel, som med hundraprocentig visshet låter en identifiera artens ägg, är synbarligen ogörligt. Från grönsångaren, den enda skandinaviska art, vars bo kan förväxlas med nordsångarens, äro äggen dock tydligt skilda. Den förra, Ph. sibilatrix, har mörkt, svartaktigt brunvioletta, tämligen väl avgränsade fläckar strödda över äggets vita yta, vanligen ganska tätt. Nordsångaren har relativt glesa, väl markerade, ibland rätt stora, mättat rödbruna fläckar, blandade med ljust, gråaktigt manganfärgade, liksom utflytande fläckar och små fina prickar, hos flertalet kullar tydligen mer jämnt fördelade över ytan än hos lövsångaren, Ph. trochilus. Dennes fläckar torde som regel vara av mindre genomsnittlig storlek och ha större tendens att vara koncentrerade kring tjockändan av ägget. Genom vänligt tillmötesgående från såväl Sune Hederström Som Göteborgs Naturhistoriska Museum, har jag fått tillfälle att granska de enda kända svenska äggen, jämte kullar från Sydvaranger och Kolahalvön. Dessa ägg se påfallande grövre ut till dimensionerna än lövsångarens.
Medeltalet för 46 ägg (Hederströms McNeiles, Rosenius’ och Wessels samt 10 ägg i Göteb. Nat. Mus. är 16,4 X 12,8 mm, medan medeltalet för 184 ägg av lövsångare Rosenius 1926) visat sig vara 15,6X12,1 mm.
Nordsångarens ägg äro tydligen i fråga om fläckarnas grovlek och anordning i en del fall ganska lika ägg av gransångaren (jfr Hederström), Ph. collybita, men dessas fläckar äro som regel mörkare brunvioletta. Gransångarens äro också något mindre, i medeltal 15,3 X 12,1 mm (59 ägg, enl. Rosenius 1926).
Antalet ägg per kull är i Skandinavien som regel 6 - 7. Av 15 kullar, för vilka säkra uppgifter finnas (L. Andersson, Collett, Hederström, McNeile, Rosenius, Swanberg Wessel) ha 9 st., d. v. s. 60 % bestått av 6 ägg eller ungar, de övriga 6 ha haft 7 ägg.
När Collett 1876 fann nordsångaren i Sydvaranger, skötos sex exemplar, och samtliga dessa hade magsäcken fullproppad med myggor. Av detta drog Collett (1877: 12) slutsatsen, att födan om sommaren så gott som uteslutande skulle utgöras av mygg. Ett par år senare skötos återigen ett par exemplar. Även dessa hade mygg i magsäcken men båda hade dessutom larver av »en Cidaria» eller någon annan mätarlarv (Collett 1881: 263). Palmén (1887: 265) fann hos flyttande nordsångare på Tjuktjerhalvön den 13 juni 1879 »skalbaggar och larver». Och Meves (1871: 757) såg, hur nordsångare den 8 augusti 1869 - den gången ny för Europa! plockade »stora gröna larver» av någon stekel (»Tenthredo eller Lophrys»).
Ungarna i boet på Nuolja fingo vid 8 - 11 dagars ålder alldeles övervägande gröna larver, vilka jag ej är kompetent att bestämma men som till typen liknade fjällbjörkmätarens, Oporinia autumnata, larv. Någon gång serverades någon småfjäril, ofta hände det att honan, då hon var färdig med en ordinarie matning med larver, snappade åt sig en mygga i luften och stoppade den direkt i gapet på en unge. Detta gick då impulsivt blixtsnabbt, liksom i förbifarten, då hon skulle ge sig iväg. Det såg alltid lika lustigt ut - det var så markerat något som låg utanför det ordinarie programmet.
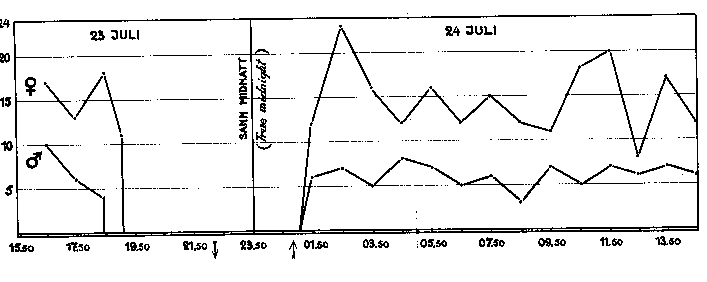
Fig. 10. Nordsångare, antal matningsbesök vid boet, per timme, 23 - 24
juli, då ungarna voro 8 - 9 dagar gamla. Punkterna markera antalet tinder
närmast föregående 60 min. Emedan solen år i undre kulminationen kl. 23.51,
är denna tidpunkt sann midnatt och timindelningen har gjorts därefter. grafik
Holger Johansson. (![]() Phylloscopus
borealis, feeding rate by e and 2, 23-24 July 1952, when the young were 8-9 days
old. The points indicate the number of feeding trips to nest in preceding 60
minutes. True knidnight is here made the basis of division in hours.)
Phylloscopus
borealis, feeding rate by e and 2, 23-24 July 1952, when the young were 8-9 days
old. The points indicate the number of feeding trips to nest in preceding 60
minutes. True knidnight is here made the basis of division in hours.)
För att få en inblick i hur ett arbetsdygn förlöper vid nordsångarens bo, placerade jag ett gömsle i närheten. Först på relativt långt avstånd, varpå det senare successivt flyttades närmare. När jag installerat mig den 23.7. kl. 15.15, kommo båda fåglarna med mat redan efter ett par minuter, men för att få en helt pålitlig och störningsfri observationsserie började jag anteckningarna först kl. 15.35.
De fortsattes oavbrutet i 24 timmar. Någon störning från människor behövde icke befaras. När honan under timmarna före midnatt sov i boet. kunde jag utan att störa, henne krypa ut bakvägen för att sträcka på benen under några minuter. Fotografering fick under denna observationsperiod ej alls förekomma. Under denna jämnt dygnslånga period kom hane med mat 104 gånger, hona 281 gånger. hane stod alltså för 26% av besöken, honon för 74%. (Se diagram, fig. 10).
Detta var alltså något om matnings – frekvensen. Gäller det emellertid att jämföra hanens totala dags -kvantitet föda med honans, blir saken svårare. Hanen kom vanligen med påtagligt större bördor än honan. Man kan absolut ej påstå, att hanen stod för blott 26 % av den totala matkvantiteten under ifrågavarande 24 timmar.
Ett stöd för detta är det förhållandet, att hane stod för borttransporten av 36 av de spillningsbollar, ungarna levererade (se nedan).
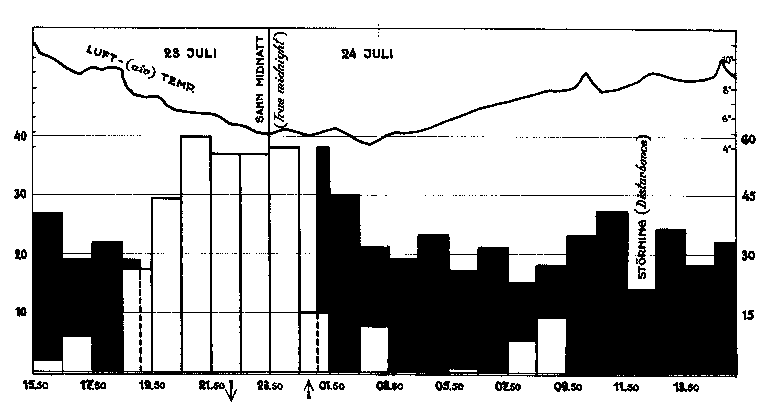
Fig. 11. Matningsintensitet och – ruvning - hos nordsångaren den 23 -
24 juli 1952. De svarta staplarna ange sammanlagt antal matningsbesök vid boet
av hane och hona. För den del av avbrotts - resp. begynnelsetimmen, då matning
pågått, har det registrerade antalet matningsbesök räknats om till den
stapelhöjd, som heltimme med samma matningsintensitet, skulle medfört. Vita
staplar ange summa antal minuter, som honan legat på boet för att värma
ungarna. Temperaturkurvan är från Abisko naturvetenskapliga station. Någon
direkt relation mellan rådande lufttemperatur och fåglarnas dygnsrytm kan ej
utläsas. (![]() Phylloscopus borealis, feeding and brooding rhythm 23 - 24 July when
young were 8 - 9 days old. Black indicates total numbers of feeding trips by
male and female, counted together. As the diagram indicates the feeding
intensity figures for cessation and initial hours have been converted into
figures that correspond with a full hour's feeding in the same rhythm. White
blocks indicate brooding by in minutes per hour. Correlation between air
temperature and activity rhythm is not direct nor absolute.)
Phylloscopus borealis, feeding and brooding rhythm 23 - 24 July when
young were 8 - 9 days old. Black indicates total numbers of feeding trips by
male and female, counted together. As the diagram indicates the feeding
intensity figures for cessation and initial hours have been converted into
figures that correspond with a full hour's feeding in the same rhythm. White
blocks indicate brooding by in minutes per hour. Correlation between air
temperature and activity rhythm is not direct nor absolute.)
Eftersom han infann sig till boet endast för matning och ej dröjde kvar där mer än nödvändigt, innebära de trettiosex procenten av renhållningen, att han genom större bördor mat, fördelade till större antal ungar per besök, stimulerat proportionsvis flera ungar till »tömningsreflex» under hans besök vid boet.
Fåglarnas dygnsrytm i norra Lappland, där det är ljust dygnet runt, är alltid ett intressant kapitel, vars djupare orsakssammanhang till väsentliga delar är okänt. Ett isolerat dygn hos nordsångaren i slutet av juli bringar oss inte närmare gåtans lösning men må anföras som en bit i problemets stora läggspel.
Översiktligt åskådliggöres de gamla fåglarnas dygnsrytm i fråga om vården av ungarna i diagrammet fig. 11. Eftersom fåglarna inte gå efter vår standardiserade tidsangivelse, har timindelningen på diagrammet gjorts i förhållande till solens undre kulmination, som inträffade klockan 23.51. Eftersom diagrammet skall i görligaste mån åskådliggöra rytmen i fåglarnas beteende, har båda makarnas matningsbesök sammanräknats och för avbrotts - respektive begynnelsetimmen räknats om till den stapelhöjd, som motsvarar den siffra, man skulle fått om den registreringstimmens matnings rytm varat under en hel timme i stället för under 35 respektive 28 minuter.
Man finner av diagrammet att matningsrytmen har varit tämligen jämn under hela aktivitetsperioden. Denna har för hanen varat till kl. 18.45 och börjat åter kl. 01.23. Honan slutade mata 40 minuter senare men började mata exakt samtidigt med hanen Hon startade från boet 2 minuter före matningen och räknar man samma förberedelsetid för hanen, får man resultatet, att matnings aktiviteten för hanen varade 17 tim. 25 min., för honan 18 tim. 05 min.
En markerad topp i aktiviteten ser man dock, som man hade anledning att vänta, i begynnelsen på morgonen, d. v. s. omedelbart efter soluppgången. En antydan till ökning av rytmen ser man också ungefär kl. 10 - 14, men om denna är signifikativ, är mera osäkert.
Här har med avsikt talats om aktivitetsperiodens längd, ej vilo eller sovperiodens, som vissa författare ibland kalla den. Den period om natten, då själva matningsdriften var avkopplad, utnyttjades inte helt till vila. Honan var under natten borta från boet fyra gånger om resp. 16, 5, 5 och 19 minuter. Det är av intresse att konstatera, att hon vid dessa utfärder ej medförde mat tillbaka. Matningen uteblev alltså inte enbart genom att den ersattes med sömn utan den var avkopplad. Även inom aktiviteten, så att säga.
Av intresse är också, att när honan under dessa nattliga utflykter avlägsnade sig, lågo ungarna hoptryckta i djup slummer både när hon lämnade dem och när hon återvände. Det var något helt annat än under dagen, då föräldrarnas ankomst varje gång utlöste en ivrig aktivitet hos ungarna. Dessa voro alltså »helt inne i rytmen».
Största intresset har emellertid aktivitetens relation till solens upp respektive nedgång. Sistnämnda värden för ifrågavarande natt i Abisko har Uppsala Observatorium beredvilligt lämnat. Med solnedgången kl. 22.32 och uppgåneren 01.10 har
Hanens matningsaktivitet upphört 227 min. före solnedgången och börjat 11 min efter soluppgången.
Honans matningsaktivitet upphört 187 min före solnedgången och börjat 11 min efter soluppgången.
Detta är i god överensstämmelse med vad vi tidigare veta om den allmänna tendensen i trakter vid eller ovan polcirkeln. Se framför allt Palmgren (1935) och Frantz (1948). Det är inte ovanligt, att fåglar i norra Lappland börja den nya dagens aktivitet redan en halv eller hel timme före tiden för solens undre kulmination (exempel betr. trastar ha lämnats i Fauna och Flora 1951: 117-119).
Med andra ord: det är inte ett visst ljusvärde som både direkt startar och avbryter dagens aktivitet om sommaren i höga Norden.
Siffrorna för nordsångaren under natten den 23 - 24 juli äro dock ej svårförståeliga, eftersom aktiviteten började först strax efter soluppgången, alltså medan ljusmängden höll på att stiga. Låt vara att ljuset ökades mycket långsamt eftersom solen under natten gick i en rätt flack båge under horisonten.
Att sedan aktiviteten avbröts så tidigt, att hanen fick vila 6 tim. 37 min., honan 5 tim. 57 min. skulle ju kunna förklaras som en följd av att tidpunkten för avbrottet står i relation till start - tidpunkt och sömnbehov. Även om båda dessa faktorer kunna ändras såväl med dagens längd som med häckningsstadiet.
Värmningen av ungarna sköttes av honan ensam och var i huvudsak inskränkt till nattens viloperiod med smärre ruvningspass under morgon och eftermiddag (se diagram, fig. 11).
Det var f. ö. nästan förbluffande att se hela den gamla fågeln försvinna in i bokulan, där redan 7 stycken 8 - 9 dagar gamla ungar lågo samlade. Hennes huvud stack dock fram i ingångsöppningen, och när hon sov, vilade det mot boets tröskel ungefär som då en människa i en tågkupé sover med huvudet vilande mot bordsskivan.
Renhållningen sköttes på det perfekta sätt, som är vanligt bland tättingar. Ungarnas »tömning» sker som resultatet av en betingad reflex i samband med matningen. D. v. s. om matsmältningen hos vederbörande unge nått ett sådant stadium, att det finns en färdig produkt att avlämna, utlöser - enligt min uppfattning – ungens sväljningsrörelser även »tömningsreflexen». Ungen gör då helomvändning, backar till boöppningen, reser sig på delvis sträckta ben, så att stjärten kommer i vädret, och pressar så fram spillningsbollen, som kommer ut omgiven av sin ytterst tunna men sega hinna. Den gamla griper så bollen, redan innan denna hunnit falla, varigenom boet ej blir förorenat (se fig. 8 o. 9). Det händer emellertid också en och annan gång, att matsmältningen nått ett sådant stadium, att ungen redan före den gamlas ankomst sitter i beredskapsställning med stjärten i boöppningen.
Ibland ägnade sig honan åt en annan sysselsättning med hygieniskt syfte, som är en vana, allmänt utbredd bland tättingar. Hon bearbetade då bobalens botten med näbben. Förfarandet motsvarar tydligen i viss mån en husmoders åtgärder vid bäddning. Ibland ser man, hur honan därvid sväljer något eller kastar ut något skräp. Denna instinkthandling är ännu ej helt förklarad, men diskussion därom torde vara onödig på denna plats. Den har fått ett frågetecken även i Miriam Rotschilds och Theresa Calys (1952:252) nyutkomna bok om fåglarnas parasiter alltifrån bakterier till gökar.
Om tiden för avflyttningen ha vi inga direkta iakttagelser i vårt land.
För Sydvaranger säger emellertid Schanning (1907), att höstflyttningen börjar strax efter det att ungarna blivit flygfärdiga, och att de sista försvunnit i senare hälften av augusti. Vid Melkefos anträffade han den 13.8.1906 en familj av hane, hona + 7 ungar, »øiensynlig på sydtraek».
För östra Finland i jämnhöjd med Torneå finner man i en tabell hos Lethonen (1943: 46), att nordsångaren 1942 skulle ha avflyttat den 11 augusti.
Slutligen kan nämnas, att Meves (1871) den 8 - 9 augusti 1869 påträffade åtskilliga familjer av nordsångare 110 - 200 km söder om Arkangelsk. Men detta är ju långt från Sverige.
Alla dessa tre uppgifter från så vitt skilda håll ge dock samstämmigt resultat, d.v.s. att nordsångarna sannolikt flytta från sina tillhåll i Skandinavien huvudsakligen i första hälften av augusti meddeförskjutningar in i senare hälften av augusti, som givetvis stundom måste bli följden av sena kullar o. s. v.
Och i sin handbok rörande östra Kinas fåglar säger La Touche (1925 - 30), att nordsångaren »är en av de tidigaste höstflyttarna», som i nordöstra Kina inträffar från mitten av augusti och visar sig i sydöstra Kina »så tidigt som under de sista dagarna av augusti». Detta gäller naturligtvis inte våra individer, men är betecknande för artens egenheter.
Nordsångaren hör till våra märkligaste flyttfåglar, och några få ord må därför sägas om vägar och vintertillhåll.
Om vår skandinaviska nordsångarpopulations vägar veta vi visserligen intet, men om teorierna är man i allmänhet överens. Förhållandet är det, att vi inte känna något enda fynd av fågeln från det egentliga Indien, där dock så många ornitologer varit verksamma, eller från Asien väster därom. Men från Bortre Indien, Filippinerna, Borneo o. s. v. och så långt ute i den malajiska arkipelagen som på Flores och Timor har man en mängd fynd av nordsångare. Man drager därför den slutsatsen att de skandinaviska fåglarna flytta genom Sibirien, östra Mongoliet och Mandsjuriet, undvikande de väldiga ökenområdena i Sinkiang och Gobi, för att genom östra Kina passera ned till vintertillhållen (Tugarinow 1931: 60, Ticehurst 1938: 126).
Det förhåller sig sannolikt så, att nordsångarens urhem legat i östra Sibirien, den har under tidernas lopp vidgat sitt häckningsområde västerut, men arten har ej i samband därmed vidgat sitt vinteruppehållsområde utan konservativt fasthållit vid det gamla och dess begränsning. Ännu tydligare blir detta, då man jämför med artens expansion mot öster. Alaskarasen, Phylloscopus borealis kennicotti, bor på den amerikanska kontinenten men flyttar om hösten konsekvent tillbaka till Asien och tillbringar vintern allmänt på, Filippinerna, mera sparsamt i övärlden söder därom och västerut t. o. m. den malajiska halvön (Parkes & Amandon 1948).
De europeiska nordsångarnas flyttning är en enorm prestation. Till de närmaste vinterkvarteren böra de från Abisko ha i rak linje omkring 7 200 km, till de avlägsnaste bortåt 11000 km, men på grund av omvägen via östra Kina torde vägen vara åtskilligt längre.
Det är mig angeläget att här tacka dem, som hjälpt mig: Svenska Turistföreningen, som gav mig det angenäma uppdrag, som resulterade i ett bo av nordsångaren, häradsskrivare Björn Holm, som introducerade mig på Nuolja, tandläkare L. Broberg, förlagschef K. Curry-Lindahl och skogsinspektor H. Viberg, vilka med stor energi deltog i genomkammandet av biotoperna vid Vittangijärvi, samt dem, som lånat ut nordsångarägg, professor 0. Nybelin och amanuens V. Fontaine vid Göteborgs Museum och överjägmästare S. Hederström. Till de övriga, som på annat sätt varit till hjälp, riktas här också ett tack!
BENT, A. C. 1949. Life histories of north American thrushes, kinglets, and their allies. - U. S. Nat. Mus. Bull. 196.
BERG, L. S. 1950. Natural Regions of the U. S. S. R. - New York.
BERGMAN, S. 1935. Zur Kenntnis nordostasiatischer Vögel. - Stockholm.
BLAIR, H. M. S. 1936. Birds of East Finmark. - Ibis (13) 6:429-459.
Carpelan J. 1927. Beitrag zur Kenntnis von dem Vorkommen und der Brutbiologie des nordischen Laubsängers im nördlichsten Finnland. - Beitr. Fortpflbiol. Vögel 3:145-147.
COLLETT, R. 1877. Mindre medd. vedr. Norges Fuglefauna i 1873-76. – Nyt
Mag. Naturvid. 23 B: 85 - 225.
- 1881. Mindre medel. vedr. Norges fuglefauna i 1877 - 80. - Nyt Mag.
Naturvid.`6 B: 2.54-394,
- 1893 - 94 Mindre medel. vedr. Norges fuglefanna 1881-92. - Nyt Mag. Natur vid.
35 B: 1-387.
Dixon J. 8. 1938. Birds and mammals of Mount MeKinley Nat. Park, Alaska. Nat. Park- Serv. Faunal Ser., No. 3. (Cit. av Tucker 1949).
Finnilä 1918. Über d. Vorkommen d. nordischen Laubsängers (Phylloscopus borealis Blas) im finnischen Lappland. - Medd. Soc Fauna et Flora Fenn. 44:14-19.
Franz J. 1948. Jahres- und Tagesrythmus einiger Vögel in Nordfinnland. - Zeitschr. Tierpsych. 6: 309-329.
HARTERT, E. & Stenbacher F. 1932-38. Die Vögel der paläarktischen Fauna. Ergräzungsband - Berlin.
HEDERSTRÖM, 8. 1944. Om nordsångarens förekomst i Sverige. - Fauna o. Flora
39: 46.
HOLM, B. 1950. Om fågelfaunan i Norrbottens län. - Norrb. Biol. Fören. småskr.:27-103.
1952. Fågeliakttagelser från Norrbottens län. - Fauna o. Flora 47: 1-5.
HORTLING, 1. 1929-31. Ornitologisk handbok. - Helsingfors.
JOUARD, H. 1935. Sur la reproduction de nos Pouillots. - Alauda 7: 127-130.
Kuusisto P. 1941. Studien über die Ökologie und Tagesrythmik von Phylloscopus trochilus acredula (L.). - Acta Zool. Fenn. 31.
LA TOUCHE, J. D. D. 1925-1930. A handbook of the birds of eastern China. London.
LEHTONEN, L. 1943. Piirteilä Pohjois- ja Keski-Vienan linnustosta. (Zusammenfass.: Grundzüge d. Vogelfauna in Nord- und Mittel-Viena.) - Orn. Fenn. 20: 33 - 57.
Lundevall C-F. 1951. X:e Orn. Kongressens exkursion till Abisko. - Norrb. Biol. Fören. småskr.: 1-8. --- 1952. Birds at Abisko. - Stockholm.
LOVENSKIOLD, H. L. 1947-49. Norges fugler. - Oslo.
MAY, D. J. 1947. Observations on the territory and breeding behaviour of the Willow-Warbler. - Brit. Birds 40: 2 -Il.
McNeile, J. H. 1952. Some notes on the nesting-habits of Phylloscopus b. borealis (BLASIUS). - Stavanger Mus. småskr. zool. ser. nr 4.
Meves W. 1871. Ornithologiska iakttagelser i nordvestra Ryssland 1869. - Övers, K. V. A. förh. nr 6: 731-788.
Niethammer G. 1937. Handbuch d. deutschen Vogelkunde, vol. 1. - Leipzig.
Palmén J. A. 1887. Sibiriska ishavskustens fogelfauna. – Vegaexpeditionens vetensk. iakttagelser, bd. 5: 245-500.
PALMGREN, P. 1935. Über den Tagesrythmus der Vögel im arktischen Sommer. - Örn. Ferm. 12: 107-121.
Parkes, K. C. & AMADON, D. 1948. The winter range of the Kennicott Willow Warbler. - Condor, 50: 86-87.
RILEY, J. H. 1918. Annotated catalogue of a collection of birds made by Mr. Copley Amory Jr., in northeastern Siberia. - Proc. U. S. Nat. Mus., vol. 54: 607-626. n%
ROSENBERG, E. 1942. Nordsångaren. I HOLMSTRÖM M. E: Våra fåglar i Norden, vol. 1. - Stockholm.
ROSENIUS, P. 1926. Sveriges fåglar och fågelbon. Vol. 1. - Lund. - (und. utgivn.) Ibidem, vol. 6.
ROTSCHILD, M. & Clay T. 1952. Fleas flukes and euckoos. - London.
Rudebeck G. 1942. Om en iakttagelse av Phylloscopus frochiloides viridanus BLYTH. - Fauna o. Flora 37: 213-219.
RUTHKE, P. 1939. Zum, Gesang des Nordischen. Laubsängers. - Orn. Monatsber. 47:181.
SCHAANNING, H. TRO. L. 1907. Ostfinmarkens fuglefauna. - Bergens Mus. Aarbog 1907, nr 8: 1-97.
- 1926. Vårtraekket i Ovre Pasvik, Sorvaranger 1908, 1910 og 1911. – Norsk Ornith. For. Tidsskr. 7:160.
SJÖLANDER, D. 1946. Zoologiska Lapplandsminnen. - Sveriges Natur 37: 95-107.
STEGMANN, B. 1931. Die Vögel des dauro-mandschurischen Übergangsgebietes. - J. f. Örn. 79: 10-236.
STEINFATT, 0. 1937. Beobachtungen über das Brutleben des Waldlaubsängers. - Beitr. z. Fortpfl.-biol. Vögel 13: 182-189.
TACZANOWSKI, L. 1872. Berieht über die ornithol. Untersuchungen des DR. Dybowski. - J. f. Orn. 20: 340-366.
TICEHURST, C. B. 1938. A systematic review of the genus Phylloscopus. - London.
TREUENFELS, H. v. 1937. Beitrag zur Brutbiologie des Waldlaubsängers (Phylloscopus sibilatrix). - J. f. Orn. 85: 605.
TUCKER, B. W. 1949. Kennicott's Willow Warbler. I A. C. Bent: Life histories of north American thrushes, kinglets, and their allies. - U. 8. Nat. Mus. Bull. 196. Tugarinow A. 1931. Die Wanderungen der nordasiatischen Vögeln. – Vogelzug 2:55-60.
ULFSTRAND, S. 1953. Iakttagelser rörande nordsångaren (Phylloscopus borealis). Fauna o. Flora 48: 25-27.
WESSEL, A. B. 1905-06. Ornithologiske meddelelser fra Sydvaranger. – Tromso Mus. Aarsh. 27:20-126.
WITIIERBY, H. F. & al. 1945. The handbook of British Birds, II. Third impr. London.
On the Eversmann's Warbler (Phylloscopus borealis BLAS.)
Swedish breeding records are three: Jukkasjärvi 25 km NE of Kiruna 1916, at the Swedish - Norwegian frontier 15 km N of the Narvik railway 1940, and Nuolja NW of Abisko 1952, all these localities between 67o55' and 68o35'N. It is supposed that the Eversrnann's warbler breeds yearly in the northern part of Swedish Lapland but it is scarce and the numbers are fluctuating from one year to another.
Plumage of young, 10 days old: whole upper - parts fairly bright greyish olivegreen. Throat yellow, other underparts yellowish white.
Soft parts of adults: upper mandible brownish black with horneoloured margins. Lower mandible light horncoloured but the anterior part of its underside was black in male brownish black in female, tip horncoloured. Legs and feet pale greyish brown.
Soft parts of young, 10 days old: bill greyish black, tip horncoloured. Angles sulphureous yellow. Mouth bright orange yellow without spots.
Song of two males 1952 resembled that of Ph. bonelli or was something between the trills of a singing Ph. sibilatrix and a singing Parus palustris.
Alarm note a short dsri slowly repeated.
Scolding note dsri rapidly repeated.
Call note is as a weak form of the alarm note.
Young, 10 - 11 days old; a hyi probably the (by H. Lynes previously described) tswee-ip.
Display once observed by Ulfstrand (1953:26-27): »30 June 1951 a male was singing in accelerated tempo and suddenly ascended in a peculiar flight, with all feathers fluffed and tail erected and with batlike wing beats, to the top of a birch where the female had perched. Then coition when the song changed into a purring si-si-si. . »
Reaction of young against closely approaching intruders: 9 - 11 days old the young made diminutive »symbolic» attacks with rapidly clapping bill and clicking sounds. A hissing sound was heard. This is not to be confused with the churring note heard from the young when they are fed by parents.
Arrives in Scandinavia normally in the latter part of June (cf. p. 63). Moves from breeding haunts in August, particularly in the first half of the month. It is supposed that nestbuilding begins almost immediately after arrival of the female In Scandinavia the first egg mormally is laid in the last week of June or the first week of July.
Eggs are 6 or 7 in Scandinavia (15 authenticated clutches; 60 %/6 and 40 %/7 eggs).
Food of the nestlings. July 1952, was for the rnain part green larvae, probably of a geometrid moth, Oporinia. The nest was watched for 24 hours, 23 - 24 July. Male fed 104 times, female 281. Male brought each time a good amount of food, female frequently only small portions. Thus the total amount of food brought by male was much larger than is indicated by the male's 26 % of the feeding visits that day.
fig.
10
fig 11
As to the daily rhythm cf. diagram figs. 10 and 11.
Only female brooded the 8 - 9 days old young. Brooding was not directly correlated to air temperature (cf. fig. 10). Sunset at 22.32, sunrise 01.10. Male fed until 227 min. before sunset, began 11 min. after sunrise. Female fed until 187 min. before sunset, began 11 after sunrise. Male activity (feeding!) period lasted 17 hours 25 min. Female activity (feeding!) period lasted 18 hours 05 min
Nest sanitation was performed in the manner common among Passerines: if digestion permitted a completed faecal sac to be delivered, the young after being fed turned, backed to the nest opening, raised its cloaca by stretching its legs and delivered its faecal sac. This was grasped by the adult just as it emerged and was not alloed to drop inte the nest (fig. 8, 9). When young were 8 – 11 days old the excrements always were carried away. In 24 hours 89 faecal sacs were removed, 36 % by 64 % by female. The percentage for male is higher than its % feeding visits to the nest. This is duo to the fact that male brought larger amount of food per visit and so many times fed two nestlings instead of the female's one. When none of the young defaecated no »prodding» was seen. The common habit among Passeriformes of »digging» in the nest bottorn was sometimes seen performed by female »Digging» was followed by brooding and is thought to be a sanitary action.
It is an interesting fact that of the species of the earlier subgenus Acanthopneuste and sibilatrix and bonelli that acc. to Ticehurst have nasal hairs as or nearly as Acanthopneuste and have a song of borealis - type the 11 with known nest - building habits all do not use feathers for nest lining (contradictory notes have been given in reguloides). Probably this indicates that these species are more closely related within their own group than to the other 17 Phylloscopus of which inornatus is said to line the nest with hairs and all the 11 others with known nest - building habits use feathers.
Studies of some species rarely photographed
P O Swanberg obtained these unique photographs of the littlestudied Arctic Warbler (Phylloscopus borealis) in 1952 in Swedish Lapland which is the northwestern outpost in this bird's breeding - range. The typical race, often known as Eversmann's Warbler, nests from there right across N. Asia, being replaced by other forms in Kamchatka, the N. Pacific islands and Alaska (which is in practice the "eastern" outpost) where it is known as Kennicott's Willow Warbler. To the north throughout most of its range the species breeds as far as the tree limit; to the south in Scandinavia it has not been found nesting below the Arctic Circle, and in Russia and west Siberia its summer range does not extend further down than about 61 degrees (roughly the latitude of Shetland), but beyond the Yenisei it reaches much further south to N. Mongolia, to Korea and Japan. This lopsided breeding - distribution is more understandable when one remembers that the entire winter - quarters of the whole species are concentrated almost exclusively in Siam, Indo - China, Malaya and the Malay Archipelago (vide C. B. Ticehurst, A systematic review of the genus Phylloscopus, 1938). The continued adherence to this restricted winter range is remarkable for a species whose breeding area has become extended to the extremes of the N. Palaearctic because, as B. W. Tucker commented (in A. C. Bent, U.S. Nat. Mus. Bull. 196, p. 333), "the European birds have not adopted what might seem the natural course of migrating for the winter to Africa." The result is an extraordinary migration and in view of the direction of it it is not surprising that the species has been recorded in Britain less than 20 times, while other birds such as the northern forms of the Bluethroat (Luscinia s. svecica) and of the Willow Warbler (P. trochilus acredula), which breed in similar areas of Arctic Scandinavia but which winter in Africa, occur regularly.
C. F. Lundevall (Fair Isle Bird Obs. Bull., no. 9 (1953), p8) says that "many records in the last few years . . . point to an increase in north Finland and northern Scandinavia since 1900." Yet the exact status is not at all perfectly known. This is partly on account of the inconstant reoccupation of breeding areas, a characteristic of Arctic birds which are dependent upon the favourability of spring weather conditions and also a feature of birds on the fringe of their breeding range, and partly because the vast areas concerned are so little worked by ornithologists. Swanberg (Vår Fågelvärld (1953), 12, pp. 49-78) suggests that the species" breeds yearly in the northern part of Swedish Lapland," but he is only able to cite 3 records, including his own, and some authors do not consider the first two of these definitely confirmed.
The Arctic Warbler, though fairly adaptable, is primarily a bird of the birch forests. Arboreal in its habits - more like the Wood Warbler (P. sibilatrix) than the Willow Warbler in this respect - it keeps to the canopy much of the time, and the cock sings his monotonous but melodious song from high in the taller trees.

Plate shows the birch - covered slope where Swanberg took his photographs; a habitat similar to that in which Robert Collett (Ibis, 1886, pp. 217-223) found three nests near Matsjok in the Tana region of Norwegian Finmark, but perhaps not a really typical habitat.
Swanberg's nest was sited on a vertical bank
Plate

The dominant field - character in the plumage of greenish above and whitish below is of course the yellow superciliary stripe that can be seen so well in the following plates. 50-52


This is made more prominent by the dark green line below it through the lores and behind the eye, below which there is another short strip of yellow. The head - pattern can perhaps best be seen in this plate
Male at nest.
In fresh plumage the greater wing - coverts and, to a lesser extent, the median ones have grayish - white tips which result in two pale wing - bars, the greater of which is usually noticeable at close range. Wear causes these tips to become less distinct and those on the median coverts quickly disappear. Sometimes the tips of the greater coverts are similarly lost and this has happened to the female in these photographs; a study of plates above will show this (note the worn appearance of the female's coverts) and give an indication of the value of the bar as a fieldcharacter.
The two following photographs
are more than good enough to illustrate the diagnostic wing - formula. No other leaf - warbler has the combination of such a minute first primary, a lack of emargination on the sixth, and pale tips to the coverts.


P O Swanberg hittade av en tillfällighet en Dvärgsparv och här beskrivs det första häckningsfyndet i Sverige. Som Du kanske vet så är detta en fågel som tillhör det Ryska riket, där gränsen nu flyttats västerut.

Breding habitat, From this birch tree almoust all song was deliverd. Parningsleken skedde i bustkarna till höger. foto 1952 H Viberg
First Swedish breeding record of Little Bunting (Emberiza pusilla PALL.)
The little bunting was found breeding about 30 kilometers NE Kiruna, 68°05' \, 20°40'E of Greenwich, in July 1952.
Previously it has been recorded breeding, or found with some evidence of
breeding, in Fennoscandia in 1849 (Schrader's "Emberiza rustica", cf Palmgren
1936), 1878 (singing male probably with a nest, Colett 1881, cf Palmgren 1936
and this paper p.230), 1907 (nest, 6 eggs, Schaanning 1913), 1911 (male shot, "marking
nest", near Kiruna, Jägerskiöld & Kolthoff 1926) and 1935 (nest and probably
another two breeding pairs, Enare in Finland, Palmgren). Finally, in July 1953 a
singing male and a female were observed in the southern Varanger district,
Norway (S. Hedenström in litt.) Thus the few records are sparsely dispersed over
more than a hundred years and no conclusions can be drawn as to the possibility
of a recent or increased spread of the species.
British Museum has 2 eggs from East Finnmark, dated 22 June 1869, but in the
author's opinion there is some uncertainty as to this record, of this paper p.
229.
Observations were made all day on each of three days, 3-5 July, and 01.45-06.00 July 6th. The habitat was a birch wood above the conifer tree limit, 400 m above sea-level. At least 3 young birds were found. The date of laying of the first egg was calculated to be June 1 and calculations based on the records of Schaannong and Palmgren lead to the conclusion that egg laying normally begins in the first 3 weeks of June in northern Fennoscandia.
Song:
Male was singing frequently 3-5 July, particularly in the morning. Song began 4
July at 00.45 (clear, temp + 2° C), 5 July 00.30 (clear, temp. +5°C). It stopped
at about 20.00 July 2d. 4 July song was interrupted at 01.32 when male started
feeding the young. After a few minutes it took up singing again, then fed for a
while; in this manner it alternated singing and feeding all the day, although
singing more frequently in the morning. True midnight at 23.41.
This male had a peculiar habit of singing that is thought to be a typical habit
for the species. Indeed, no other Emberiza, known by the author, sings anything
like thismale of E. pusilla. The pleasant song was built up of several different
phrases, each phrase repeated a number of times (10-20) before the bird changed
to the next phrase. Each phrase could be varied in itself. The four principal
types of them were:
1) with opening notes like those of yellow bunting (Emberiza citrinella,),
2) with rapidly rattling notes like a series of notes in the song of the tree
pipit (Anthus trivialis),
3) resembling the ortolan bunting (Emberiza hortulana) as near in pitch as if it
were imitating (but it is not considered to be imitation!),
4) a more twittering phrase, suggestive of linnet (Carduelis cannabina) or
Lapland bunting (Calcarius lapponicus). The phrases are spelt (in the Swedish
way) p.218 where numbers correspond to the numbers above in this summary. After,
e.g., 10-20 phrases of no. 1 could follow as many of no. 2. Then no. 2 was
interrupted for a sequence of no. 3 and so on.
In full song about 10 phrases per minute were delivered. With few exceptions the
male was always singing from the top of the largest tree, a birch of 8-9 meters
(Fig. 6). In song the frontal feathers were raised.
Call note:
"kitt", high-pitched, thin. This note was used by adults when calling young.

The principal feedingground of little bunting. Larvae were collected in the low vegetation of the ground close to Vittangijärvi.
Alarm note:
"ksitt", with audible "s", recalling the common call note of song-thrush but
thinner and difficult to distinguish from that of the rustic bunting.
Display was apparently performed many times in the morning hours of July 3, 4 and 5, and also later in the day. Usually it was heard at a distance of a few meters but not seen in the dense willow-scrub. It was observed 4 July at 16.05: male in loud song, at the top of the song tree, female sneaking in the favorite willows just beneath the song-tree, male dived into the willow-scrub of female and when there his song changed into a subdued version of "the ortolan bunting-phrase", or subsong. Female took her place at a bare spot on the ground. Then male with wings extended upright and without audible notes danced on the ground in S-shaped curves in a semi-circle in front of female. Coition followed in the manner common to buntings and others. The next action was probably an abnormal one, an "inversion": female jumped up on the back of male who pressed himself to the ground, quite motionless for a few moments. After separation female perched in the scrub and preened the rump.
Young were fed by male and female with green larvae, collected mostly among
Vaccinum myrtillus and other low vegetation, sometimes in the birch trees up to
3 meters above the ground. Young birds were scattered widely in the wood. One
young bird was fed repeatedly, then the adult flew to another, sometimes 200
meters from the first one, and fed it 3-5 times in succession and so on.
Double-brooded or not ? The behaviour - song throughout the day and the frequent
displays - suggested that the birds prepared a second brood but, in spite of all
efforts, watching all the day, there was no proof of it. Portenco (1937) says:
It is supposed that Emb. pusilla, too, is double-brooded".
Injury-feigning by male resembling that of the reed - bunting was seen when a
young bird was flushed while the male was feeding it.
The paper gives a short account of other aspects of the breeding biology, general distribution, distributional history and migration, based on the literature.
Vill Du veta mer kontakta Biblioteket i Skara eller uppsök Vår Fågelvärld Årg 13 1954 Nr 4: H Viberg var med när detta hände.